
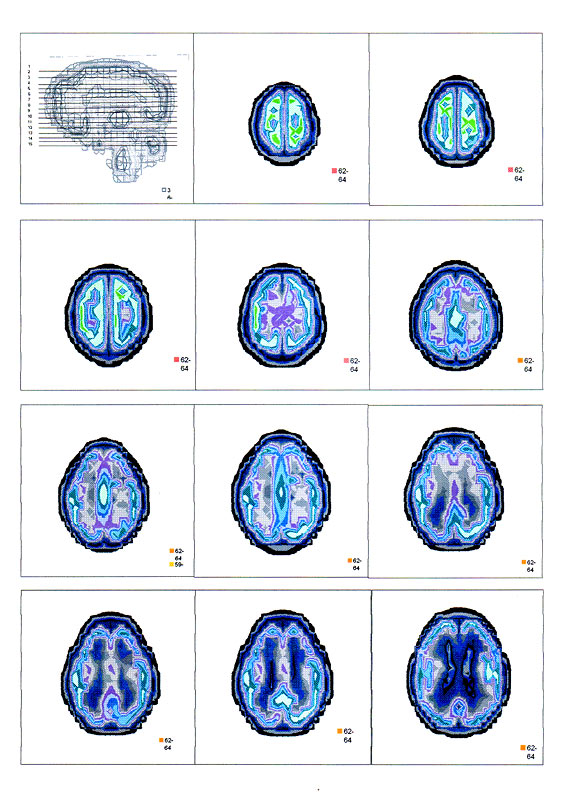
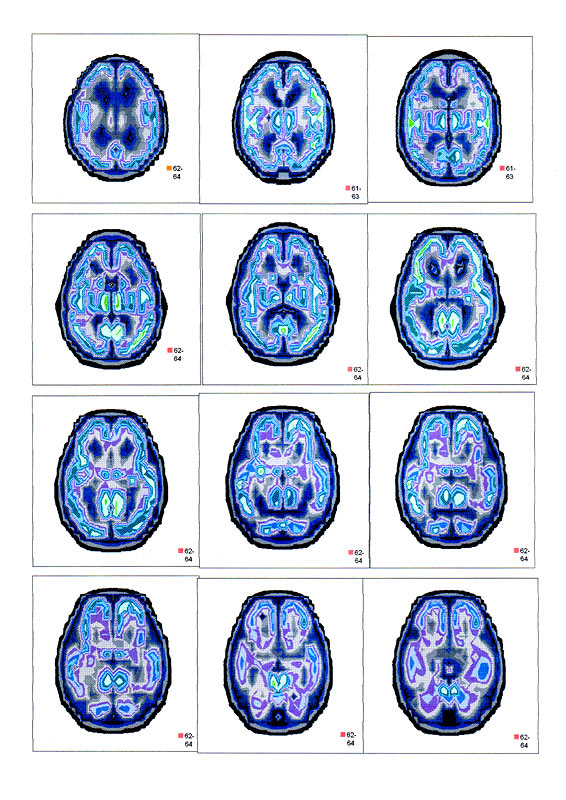
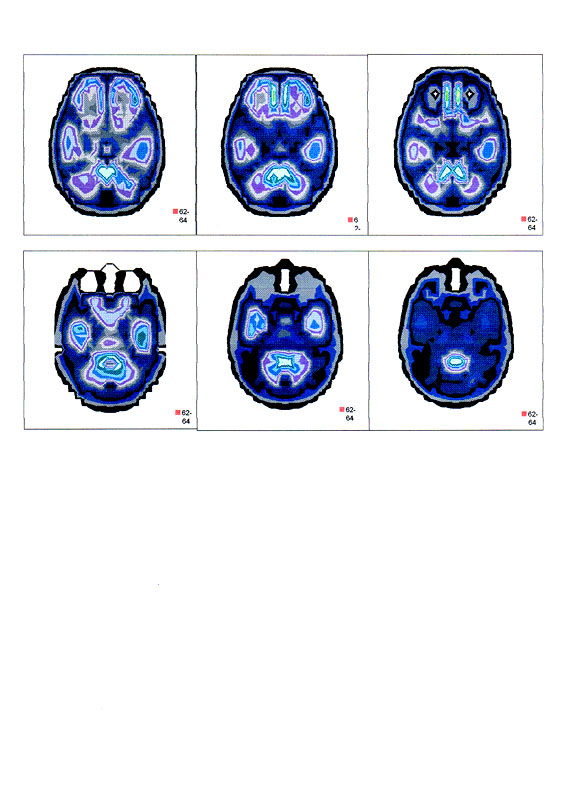
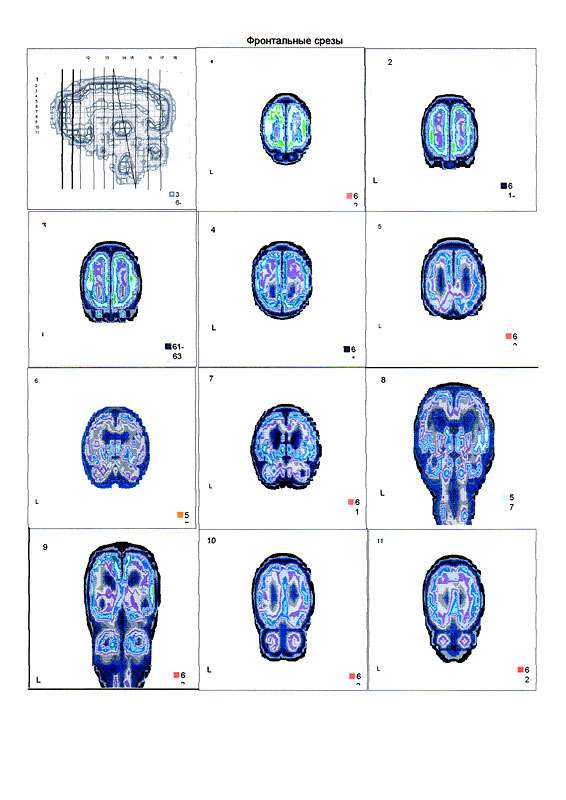
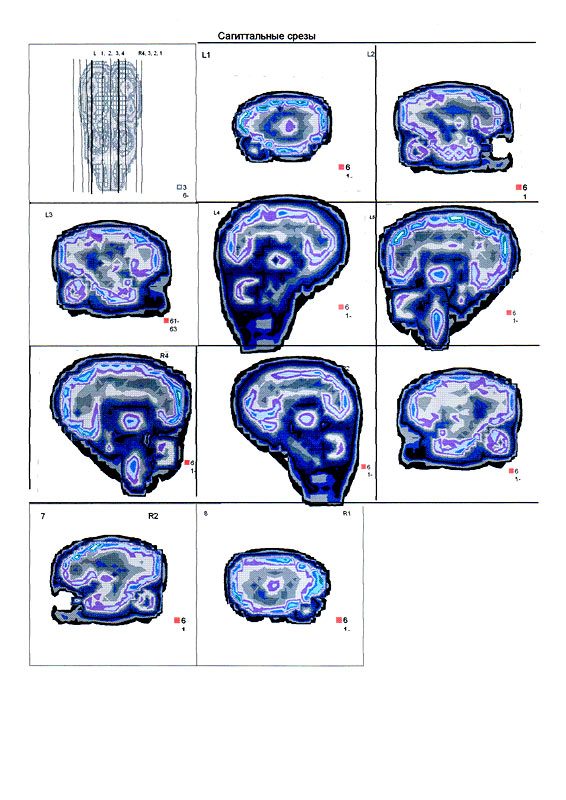
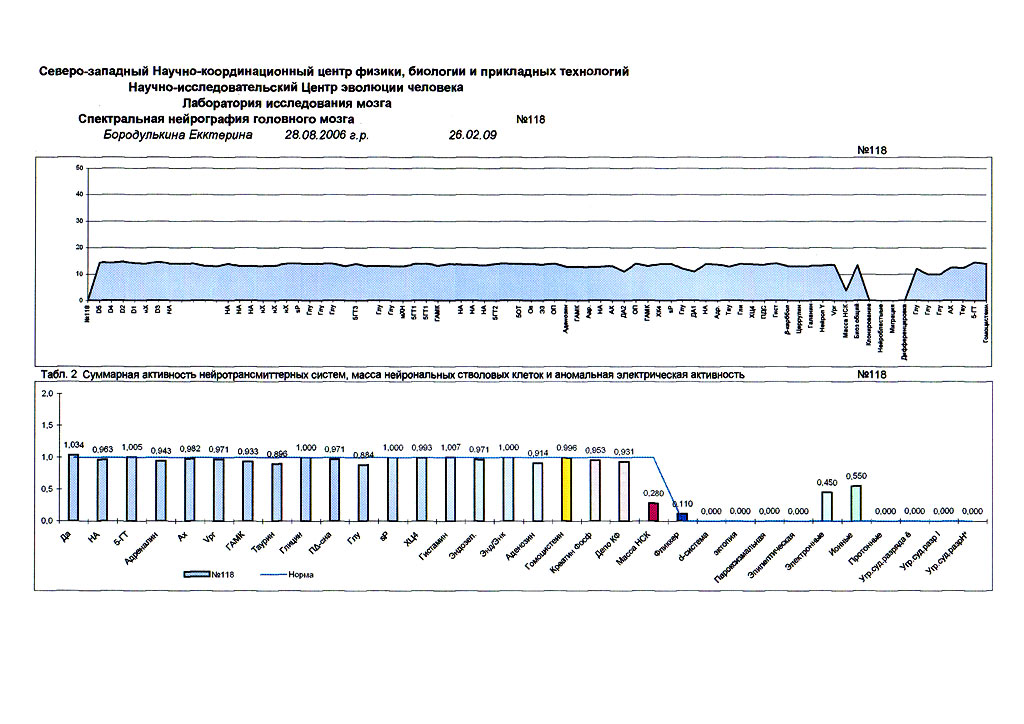
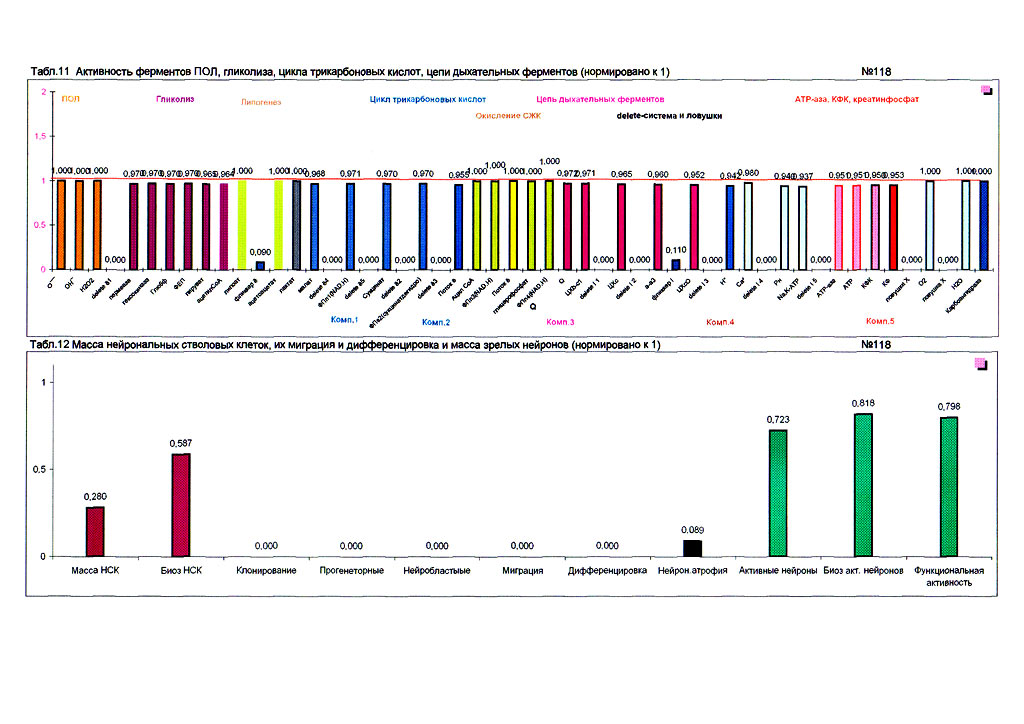
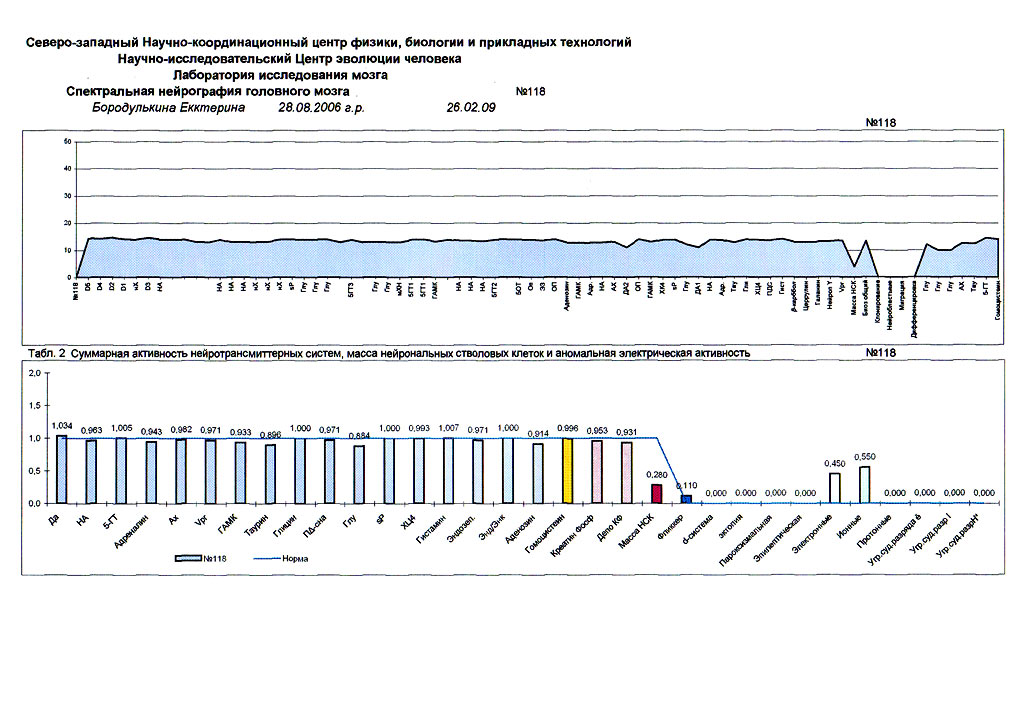
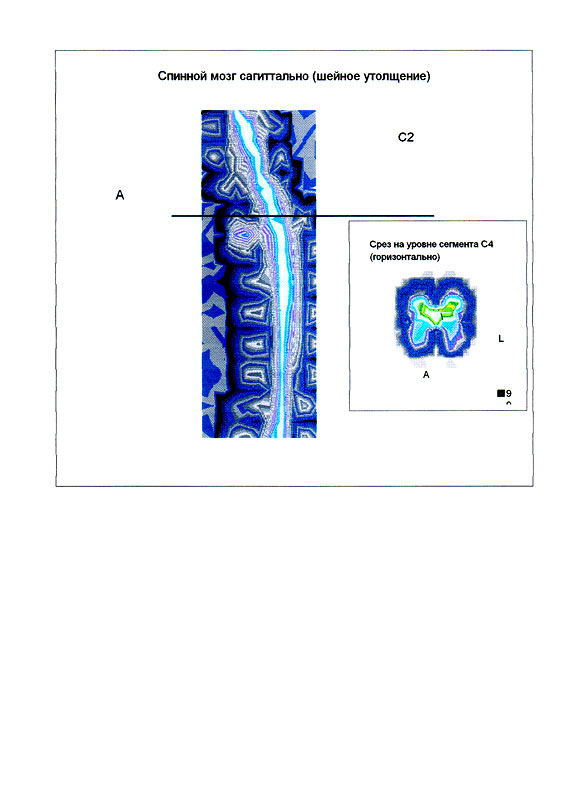
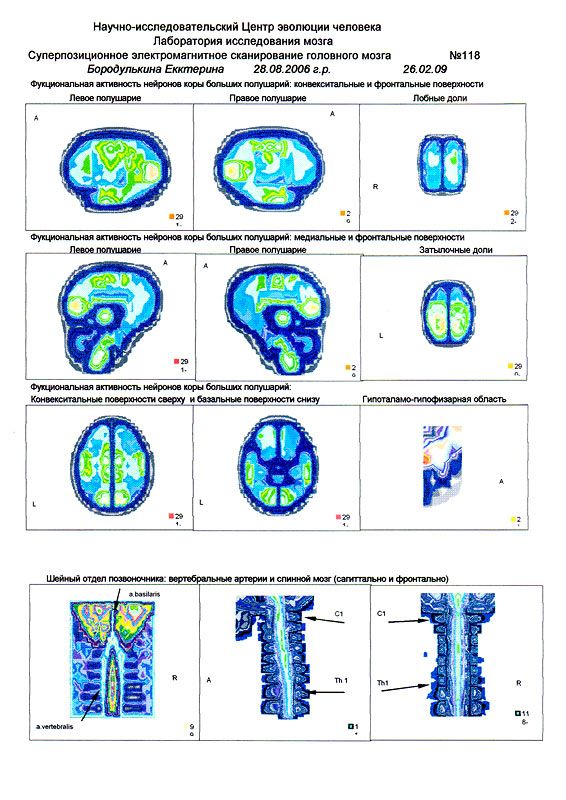

| Суперпозиционное электромагнитное сканирование головного мозга, 2 года 5 месяцев, 26.02.2009, Камынин Ю.Ф., Северо-западный научно-координационный центр физики, биологии и прикладных технологий, Научно-исследовательский центр эволюции человека, Лаборатория исследования мозга | ||
| |
||
| |
||
|
||
|
||
|
||
| |
||
| |
||
| © 2009 www.katerinab.ru | ||